
مکانیسم های مرتبط به جذب چربی و عوامل موثر بر آن در طیور
چربی های استفاده شده در تغذیه طیور از طریق مکانیسم های مختلفی هضم، جذب و مورد استفاده قرار می گیرند که به صورت مختصر در این مقاله مورد بحث قرار می گیرد.
هضم چربیها در طیور
در همه حیوانات بخش اصلی هضم و جذب چربی غذا در روده کوچک اتفاق می افتد، زیرا بخش عمده ای از آنزیم ها و ترکیبات موثر بر هضم چربیها همانند لیپاز ,کلسترول استراز و بیکربنات مترشحه از اندام های ضمیمه دستگاه گوارش به بخش ابتدایی روده کوچک ریخته می شوند. هضم چربی ها با امولسیفیه شدن آنها توسط نمکهای صفراوی مترشحه از کیسه صفرا تشدید می شود. مایع صفراوی علاوه بر نمک های سدیم و پتاسیم، اسیدهای صفراوی که عمدتا اسیدهای گلیکو کولیک و تائوروکولیک هستند حاوی فسفو لیپیدها (بخش لیستین)، رنگدانه های صفراوی بیلی روبین و بیلی وردین (محصولات نهایی کاتابولیسم هم ) کلسترول و موسین نیز میباشد. نمک های صفراوی علاوه بر امولسیونه کردن چربی ها، به واسطه فعال کردن آنزیم لیپپاز لوزالمعده ایی نقش مهمی در هضم چربیها ایفا می کنند.
کلسترول استراز که از لوزالمعده ترشح می شود، استرهای کلسترول و اسیدهای چرب را به کلسترول و اسیدهای چرب آزاد هیدرولیز می کند.
آنزیم لستیناز A نیز به صورت غیرفعال از لوزالمعده ترشح و به وسیله آنزیم تریپسین و با حضور یون کلسیم فعال می شود. هیدرولیز محدود پیوندهای استری در کربن شماره 2 لستین و تولید لیزولستین سبب اتصال لیپاز به سطح مشترک مایع و چربی سوبسترا شده و هیدرولیز تری اسیل گلیسرول را تسریع می کند. کولیپاز نیز پروتئینی است که در شیره لوزالمعده وجود داشته، به سطح مشترک املاح صفراوی، تری اسیل گلیسرول ها و آب متصل شده و تکیهگاهی با میل ترکیبی زیاد برای لیپاز به وجود میآورد. کولیپاز برای فعالیت لیپاز ضروری می باشد. لیپاز لوزالمعده برای اسیدهای چرب که در موقعیتهای 1و3 به گلیسرول متصل شدهاند به طور اختصاصی عمل کرده و این ویژگی ابتدا منجر به تولید 1و2-دی گلیسیریدها و سپس 2-منو گلیسیریدها، اسیدهای چرب و یا حتی گلیسرول خواهد شد.
چندین عامل باعث ترشح شیره پانکراس به داخل دوازدهه می گردد. در زمان ورود اسید به دوازدهه، هورمون سکرتین از مخاط روده به داخل خون ترشح شده و این هورمون باعث تحریک سلول های پانکراس گشته و آنها را وادار به ترشح مایع رقیق می نماید. مایع مزبور حاوی مقدار زیادی یون های بیکربنات و مقدار کمی آنزیم است. هورمون دیگری به نام کوله سیستوکنین ( پانکروزیمین) نیز به هنگام ورود لیپیدها و سایر مواد هضمی به داخل دوازدهه، از مخاط آن ترشح و باعث تحریک ترشح لیپاز و لستیناز از پانکراس می شود.
نتیجه نهایی هضم چربیهایی که از راه غذا و یا صفرا (فسفولیپیدها) به روده رسیدهاند شامل2-منوگلیسیریدها به عنوان فراورده اصلی گوارش تری آسیل گلیسرول ها و همچنین گلیسرول، اسیدهای چرب، کلسترول و لیزو فسفو لیپیدها می باشد.
جذب چربی ها در طیور
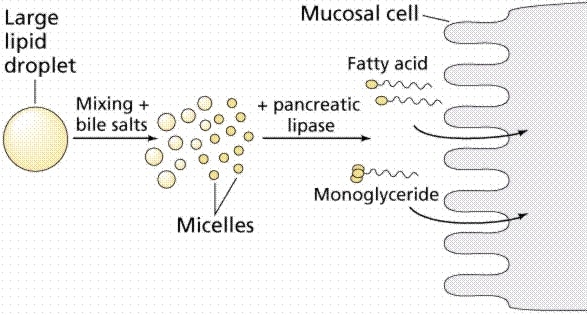
پس از هضم چربی ها در روده، بخشی از منوگلیسیریدهای آزادشده به همراه اسیدهای چرب با زنجیره کوتاه و متوسط به صورت دستنخورده جذب می شوند. با این وجود بیشتر منوگلیسیریدهای آزادشده و اسیدهای چرب غیراشباع قطبی که به تنهایی قادر به تشکیل مسیل نیستند به راحتی با نمکهای صفراوی، لستین و مولکول های کلسترول، میسل های مخلوط پایداری را تشکیل می دهند. ترکیبات موجود در ساختمان میسل های مخلوط قادر به حل مقدار قابل توجه ای از اسیدهای چرب اشباع غیر قطبی همانند اسید پالمتیک و استئاریک و ویتامین های محلول در چربی در خود هستند. به این ترتیب اسیدهای چرب و سایر مواد شبه لیپیدی در فاز آبی لوله گوارش محلول بوده و به آسانی با پرزهای روده تماس یافته و به سطح درونی مخاط روده باریک رسیده و از آنجا به وسیله انتشار غیرفعال جذب می شوند.
مکانیسم جذب اسیدهای چرب زنجیره کوتاه و متوسط که به طور مستقیم و بدون نیاز به تشکیل میسل است، برخلاف شیب غلظت بوده و به وسیله انتقال فعال وابسته به سدیم است. بیشترین میزان جذب لیپیدها در ناحیه ژژنوم روده کوچک است، به طوری که حدود 20 % درصد از لیپیدها در ابتدای ژژنوم و 60 % نیز در قسمت های میانی و انتهایی ژژنوم جذب می شود.
عوامل موثر بر هضم چربی در طیور
چربی ها و اسیدهای چرب به عنوان منابع انرژی قابل دسترس برای جوجه ها تفاوتهای زیادی با هم دارند. این تفاوت ها تحت تاثیر قابلیت هضم و جذب اسیدهای چرب از روده است، زیرا اسیدهای چرب از طریق ادرار دفع نمی شوند. بدین ترتیب با مشخص کردن انرژی کل چربی ها به وسیله بمب کالریمتر و ضرب آن در درصد قابلیت هضم آنها می توان انرژی قابل سوخت و ساز چربیها را محاسبه کرد. قابلیت هضم چربی ها در طیور توسط عوامل زیر تحت تاثیر قرار می گیرد:
- طول زنجیره اسید چرب : اسیدهای چرب با زنجیره کوتاه و متوسط بهتر جذب می شوند.
- تعدا پیوندهای 2 گانه اسیدهای چرب
- وجود یا عدم وجود پیوند استری: به طور کلی شکل منوگلیسرید اسیدهای چرب اشباع و غیر اشباع بخصوص هنگامی که در موقعیت 2 تری گلیسرید هستند بهتر از اسیدهای چرب آزاد جذب می شوند.
- سن جوجه
- نسبت اسیدچرب غیر اشباع و اشباع: اسیدهای چرب غیر اشباع و انواع منوگلیسریدها براحتی با نمکهای صفراوی میسل مخلوط تشکیل داده و این میسل ها اسیدهای چرب اشباع را به صورت محلول در می آورند. به طور کلی ترکیب اسیدهای چرب غیر اشباع و اشباع باعث اثر همکوشی می شود و این نسبت 4 به 1 اعلام شده است.
- ترکیب جیره: سطوح بالای فیبر غیر قابل هضم در جیره تاثیر منفی بر قابلیت هضم چربی ها دارد.
- عوامل روده ای: pH مواد هضمی می تواند هضم چربی ها را تحت تاثیر قرار دهد، زیرا شرایط اسیدی حلالیت میسل ها را کاهش میدهد.
انتقال و ذخیره چربیها در طیور
مشخص شده که گلیسرول آزاد درون روده و اسیدهای چرب کمتر از 10 تا 12 کربن پس از جذب به طور مستقیم وارد خون وریدی باب شده و به کبد می روند. در سلولهای مخاطی روده 2-منواسیل گلیسرول با شکل فعال اسیدهای چرب بلند زنجیر (آسیل کوآنزیم) پیوند استری تشکیل داده و تری گلیسیریدها را می سازند. همچنین لیزوفسفولیپیدها و قسمت زیادی از کلسترول جذبشده نیز با آسیل کوآنزیم آ ترکیب شده و فسفو لیپیدهای مربوطه و استرهای کلسترول را به وجود میآورند. به این ترتیب تری آسیل گلیسرول های سنتز شده به همراه فسفولیپیدها (به خصوص لستین)، استرهای کلسترول و لیپوپروتئینها تشکیل کیلومیکرون ها را می دهند.
در طیور کیلومیکرون ها پس از تشکیل، مستقیماً جذب ورید کبدی شده و بعد از ورود به جریان خون به کبد منتقل می شوند. در طیور بر خلاف پستانداران کبد محل اصلی متابولیسم چربی میباشد. در این بافت چربی های جذبشده و همچنین چربی سنتز شده از قندهای اضافی، ذخیره و سپس چربی های جدید که عمدتا به صورت لیپو پروتئین با وزن مخصوص خیلی کم هستند، از طریق جریان خون به بافتهای مختلف از جمله بافت چربی منتقل می شوند. در بافت نیز به دلیل اینکه فقط اسیدهای چرب آزاد می توانند از غشاهای چربی عبور کنند، تری گلیسریدها توسط آنزیم لیپوپروتئین لیپاز هیدرولیز شده و اسیدهای چرب حاصله وارد سلول می شوند.